The AI Picbreeder Experiment
Can AI agents be creative? It's a question on the lips and fingertips of many. Sure, AI is the new medium, but could it ever be the one making the message? We ask whether the agents we use today as tools might be re-purposed to spontaneously speak among themselves. The end result would be a model organism of cultural production.
What should we ask of the agents to make this real? The tension is that we cannot ask for anything at all—it has to be left up to them—yet modern agents demand our asking by design. They are trained, evaluated, and orchestrated as goal-following entities. But creativity elides planning, and the end products of cultural processes are not conceived of at the outset but forged in serendipity. This is true not only of the arts, but of math and science as well, where the discovery of new questions is at least as important as their resolution.
To this end, we envision agents capable of intentional aimless wandering—agents that can surprise themselves, and in response throw their plans and preconceptions out the window in pursuit of the unexpected.
To study whether AI systems might have this capacity, we turn to a minimal substrate where we know it to be possible—even necessary. Picbreeder



Before frontier models allowed us to speak into existence a vast distribution of possible images, Picbreeder had users breed images by representing them indirectly.
Each image in Picbreeder is encoded as a Compositional Pattern-Producing Network (CPPN)
In practice, we pass all the input coordinates through the network at once. Each node in the network has some grid-shaped activation we can render as a greyscale image, and at the network's output, three such outputs are combined to produce a single color image. In the tool below, you can see how changing the weights or activation functions of various nodes and edges changes the intermediary activations and final output of a CPPN:
Playing with the weights for a moment, you get the sense that building target images by building CPPNs from scratch out of nodes and edges would be a thankless endeavor. Instead, in Picbreeder, images are evolved. This is done interactively with the user in the loop: from a grid of random initial CPPNs (which generally look like smooth blobs or gradients), a user selects one or several which they'd like to grow further.
These individuals (each little brain containing a single picture) are then bred together and mutated to produce a new set of candidate images, and the process repeats.
Mutation and crossover operations are drawn from the Neuroevolution of Augmenting Topologies (NEAT) algorithm
In this way, images are created not by design but by a prolonged process of natural selection. And as the human in this loop, it's nearly impossible to plan for a particular long-term goal: for example to imagine an image in advance then successfully breed it. Not within this interaction scheme, not on any reasonable kind of timescale, at least. But Picbreeder shows us that these kinds of achievements can be attained anyway if the process of interactive evolution is distributed across a large number of users. Here, the artifacts materialize by accident, appearing to the user as objectives-in-retrospect.
In other words, the Picbreeder users played with the system, wandering, and generated art not by conceiving it against marble or canvas, but by catching it like a fish from a stream. In the AI Picbreeder experiment, we ask: can AI catch similar fish? And if it can, then how does it achieve this? Then we can try to find any parts of its looking at and moving in the water that matter to its success.
To get at these questions, we conceive of a simplified interaction loop with the Picbreeder website, automating users with large vision-language models. In this loop, we sample from an ever-growing shared archive of publications, and feed these snapshots to various parallel VLM agents. These agents can select an image to "branch", breeding its offspring across generations, then naming and publishing their favorite child. Intermittently, agents are also asked to evaluate the quality and novelty of such snapshots from the archive. Future samples are drawn partially at random, and partially according to recency of publication, mean ratings from past agents, and branching popularity.

This resembles Evolution through Large Models
Such an experiment in open-endedness cannot have a pre-defined objective. But while developing the system, examining its outputs, and reconstructing the phylogeny of the human archive, we concluded that qualitatively, the human archive has a certain je ne sais quoi. There is a kind of discernment among the forms there, a refinement lacking from some of the AI galleries, as if the AI agents never got over the initial spark of resemblance to find something better yet.
Indeed the agents' work projects a certain je sais exactly what. They often get sucked into their own attractors. Where the humans seem to capitalize on luck and imbue it with attention to create something refined, the AI often seems to experience it more passively, if perceptively. For what it's worth, though, the attractors that show up are different enough between seeds and hyperparameters to make repeatedly running the system a relatively entertaining endeavor.
Have a look at the galleries by clicking the thumbnails below:

With some idea of the shape of this qualitative difference in mind, we architect some metrics to express it numerically. In sum, we try to measure how visually different images in the archive look, how much they look similar to a predefined set of objects, and—beyond this predefined set—the diversity among the things they depict (i.e. in terms of the captions a model would generate for them).




We also develop a choice set of knobs for the system. We're inspired by the idea that serendipity requires chance, a kind of local narrative flow, and a broader personal context; and design interventions intended to serve as simplified computational models of these effects. In particular, we inject random noise into the agents' decision making process, we play with their memories—erasing everything but the present moment or overloading them with a full view of everything they've ever seen—and we seed them with simple personality traits intended to indirectly influence their interaction with the system.

We find that the historical human archive tends to dominate VLM runs in terms of our evaluation metrics. We additionally measure the tree balance of generated phylogenies to gain a sense of the extent to which agents and users fixate on a choice few prolific parents when branching. Without interventions, VLMs tend to be more discriminatory/myopic/obsessive than humans by this measure, while the random baseline—by definition—is the most balanced.
When augmenting the VLM loop with agent diversity, the system—on separate occasions—catches up in terms of semantic recall and semantic coverage. Adding agents also tends to steadily increase the degree of balance in the tree.
When we remove agents' memories, and they only have context of the current generation, they're prone to reproducing the same image ad nauseam. Even one previous generation of history is enough to break this pathology, making the agent less likely to knowingly repeat a selection. Too much memory seems to overload the agent's context and degrade its capabilities, with archives at longer memory-lengths often filled with overly abstract or repetitive, simple forms.
Injecting noise in agents' breeding selections can also—if a little crudely—push the agents out of problematic basins, at the risk of filling the archive with a greater proportion of intermittent stepping-stone slop (though interestingly, recognizable forms eventually do emerge even when agents are restricted solely to branching decisions; and indeed we can see this dial as a second-order mutation rate).
| Sweep | Setting | Semantic Recall | Visual Coverage | Semantic Coverage | Tree Balance (J¹) |
|---|---|---|---|---|---|
| Noise (ε) | 0.0 | 0.087 ±0.001 | 0.614 ±0.002 | 0.696 ±0.004 | 0.235 ±0.026 |
| 0.05 | 0.086 ±0.001 | 0.619 ±0.011 | 0.702 ±0.006 | 0.246 ±0.023 | 0.25 | 0.088 ±0.001 | 0.638 ±0.009 | 0.717 ±0.004 | 0.249 ±0.022 |
| 0.5 | 0.085 ±0.001 | 0.633 ±0.007 | 0.709 ±0.005 | 0.260 ±0.013 | |
| 0.75 | 0.084 ±0.001 | 0.639 ±0.004 | 0.706 ±0.003 | 0.303 ±0.010 | |
| 1.0 | 0.082 ±0.001 | 0.610 ±0.013 | 0.700 ±0.003 | 0.275 ±0.030 | |
| Memory (CL) | 0 | 0.082 ±0.001 | 0.527 ±0.009 | 0.632 ±0.006 | 0.305 ±0.027 |
| 1 | 0.087 ±0.001 | 0.614 ±0.002 | 0.696 ±0.004 | 0.235 ±0.026 | |
| 2 | 0.083 ±0.001 | 0.583 ±0.004 | 0.675 ±0.006 | 0.339 ±0.033 | |
| 10 | 0.079 ±0.003 | 0.512 ±0.021 | 0.661 ±0.010 | 0.331 ±0.040 | |
| 20 (full) | 0.083 ±0.001 | 0.595 ±0.010 | 0.697 ±0.003 | 0.350 ±0.018 | |
| Agents (NA) | 0 | 0.087 ±0.001 | 0.614 ±0.002 | 0.696 ±0.004 | 0.235 ±0.026 |
| 10 | 0.086 ±0.002 | 0.605 ±0.012 | 0.698 ±0.011 | 0.373 ±0.030 | |
| 100 | 0.089 ±0.001 | 0.659 ±0.006 | 0.710 ±0.009 | 0.473 ±0.022 | |
| 1000 | 0.088 ±0.001 | 0.665 ±0.004 | 0.734 ±0.013 | 0.476 ±0.013 | |
| Random | 0.080 ±0.001 | 0.612 ±0.005 | 0.692 ±0.002 | 0.540 ±0.003 | |
| Human | 0.089 | 0.681 | 0.730 | 0.363 | |
Elsewhere, it’s been argued that with respect to training large models like LLMs, open-ended search might be crucial to learning good representations
We see some evidence of that here, perhaps, in a skull form produced by AI Picbreeder—without our asking, and indeed the AI variant is plainly lesser than the human one. Sweeping the most consequential CPPN weights of this image does not explode it as in a CPPN optimized to produce such a skull via gradient descent. It remains to be seen, however, how much of this representational robustness is owing to CPPNs and NEAT, and how much to the VLMs' judgment and the Picbreeder interaction loop at large. But these experiments would seem to suggest that at least some of one potentially important emergent byproduct of open-endedness can recur in a synthetic system without its being optimized for it directly.
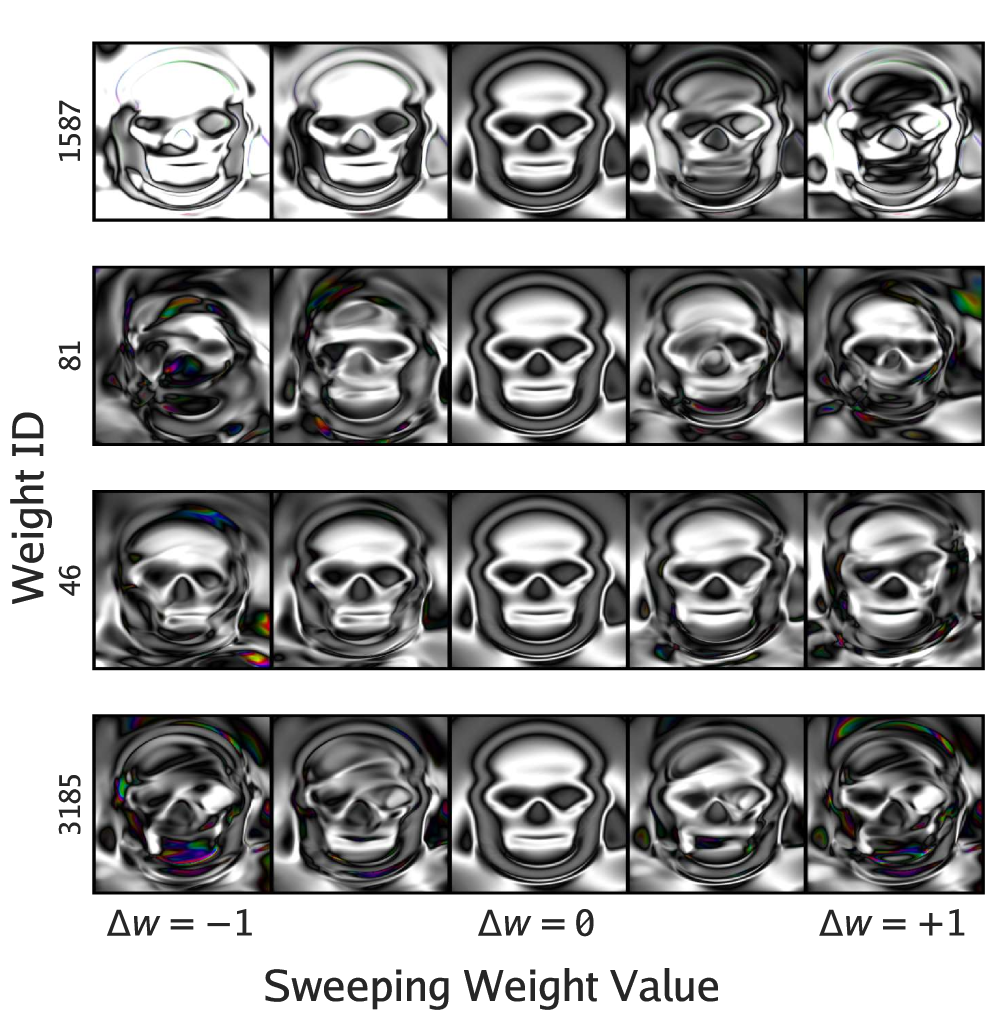 SGD-trained CPPN. Perturbing individual weights causes skull-crushing distortions—the "fractured, entangled" regime. From
SGD-trained CPPN. Perturbing individual weights causes skull-crushing distortions—the "fractured, entangled" regime. From  VLM-evolved CPPN (ours). Perturbations stay skull-like and change relatively smoothly—less fractured than SGD, but without crisp semantic factors.
VLM-evolved CPPN (ours). Perturbations stay skull-like and change relatively smoothly—less fractured than SGD, but without crisp semantic factors.
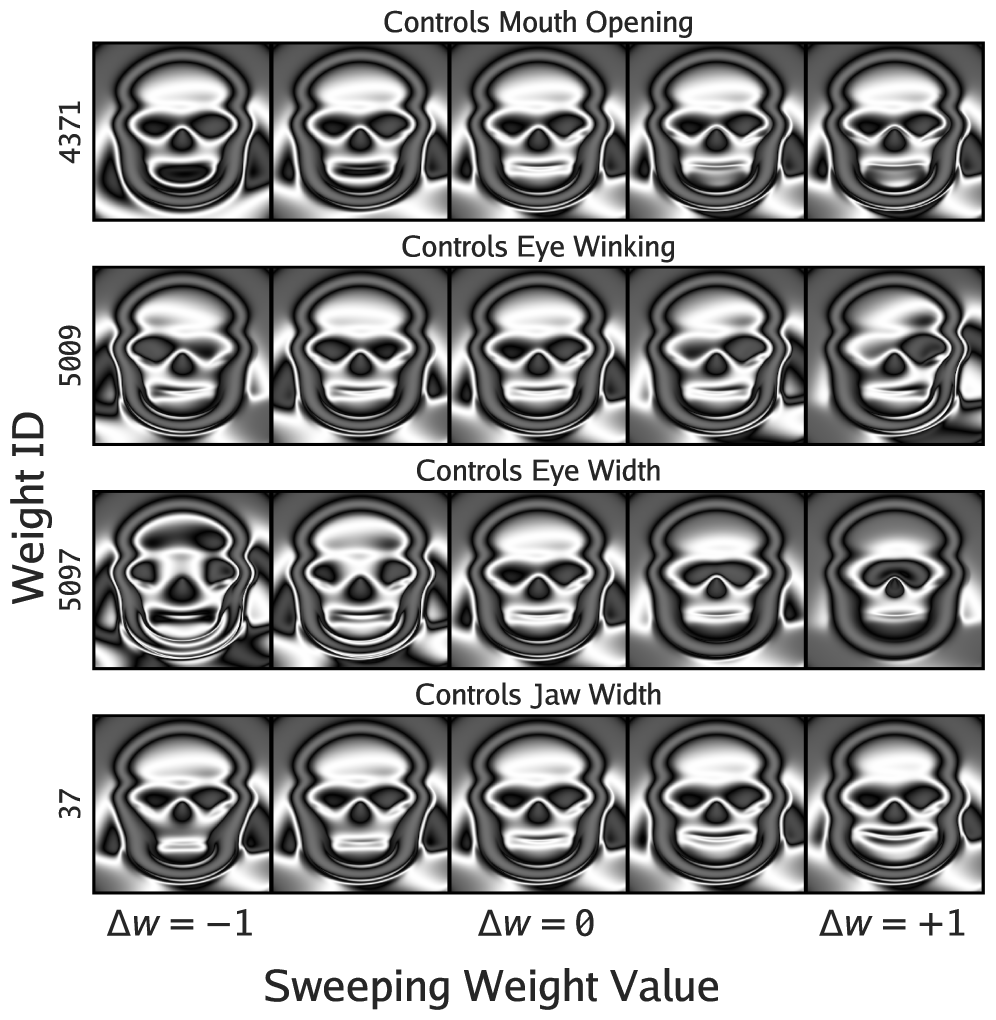 Human-evolved Picbreeder CPPN. Individual weights cleanly factor into semantic controls—"Mouth Opening", "Eye Winking", etc. From
Human-evolved Picbreeder CPPN. Individual weights cleanly factor into semantic controls—"Mouth Opening", "Eye Winking", etc. From But this doesn't mean that synthetic open-endedness is not necessarily solved. One could contend, for example, that VLM-driven Picbreeder is not engaged in a process of open-ended discovery so much as of rediscovery—of circuitously and arduously recovering its vanilla visual priors through a molasses of neural image representations. Though here we'd counter that a large part of human creativity in this space could be cast in the same light, that this is by design a space where creativity operates under and is characterized by strong constraints.
No, maybe in spite of our efforts to engineer a pipeline for collaborative design, the agents driving it are plagued by an irreconcilable rigidity. We can see traces, perhaps, of their striver pathology in the transcripts. See below, where two vehicles are arrived at via more or less winding roads; the VLMs tend to remain in the semantic space of cars in general (moving from the driver's seat, to the dashboard, to the hood, to a full profile) while humans traverse a more eclectic and unpredictable sequence, exploring eyes and (alien) faces before suddenly landing on the automobile. In this light, one could argue the VLMs show a lack of imagination, where even the apparently serendipitous leaps in semantic space are in fact predictable according to an implicit theme or predetermined objective.















We make the code and data available for follow-up questions. Though here we’ve distilled from the design of Picbreeder.org a kind of simple, bespoke evolutionary algorithm, there might be some benefit in taking a more unstructured, true-to-life approach. Our homage to the former website acts as a first step in that direction: a number of agents could be allowed to interact with the site's various pages open-endedly, browsing, searching, and jumping between sessions at their own pace.